
Int J App Pharm, Vol 17, Issue 2, 2025, 306-313Original Article
EXPLORING THE NEUROPROTECTIVE POTENTIAL OF GEISSOSPERMINE AND HARMINE ON DOPAMINE D1 RECEPTORS: AN IN SILICO STUDY
SRIRANJINI A. S., ASHSIH THAPLIYAL*, KUMUD PANT
Department of Biotechnology, Graphic Era Deemed to be University, Dehradun-248002, India
*Corresponding author: Ashsih Thapliyal; *Email: ashish.thapliyal@geu.ac.in
Received: 12 Apr 2024, Revised and Accepted: 06 Feb 2025
ABSTRACT
Objective: This study aims to explore the binding affinities and interaction mechanisms of the phytocompounds geissospermine and harmine with the Dopamine D1 receptor (D1R), providing insight into their potential as therapeutic agents for neurological disorders.
Methods: The three-dimensional structure of the Dopamine D1 receptor was obtained from the Protein Data Bank (PDB), while the structures of geissospermine and harmine were retrieved from the PubChem database. Preliminary screenings, including secondary structure validation and pharmacokinetic profiling, were conducted to assess the suitability of these compounds for docking studies. Molecular docking was performed using AutoDock Vina to predict the binding affinities and interaction profiles of geissospermine and harmine with D1R. To further validate these interactions, Molecular Dynamics (MD) simulations were conducted using GROMACS v2019.4 to assess the stability and behavior of the geissospermine-D1R complex over time. Parameters such as Root mean Square Deviation (RMSD) were analyzed to evaluate the dynamic stability of the protein-ligand interactions.
Results: Molecular docking revealed that geissospermine exhibited a higher binding affinity towards D1R with a binding energy of-8.6 kcal/mol, interacting with key residues including ILE 1009, ARG 1008, ASN 311, VAL 22, GLU 85, THR 26, ALA 57, LEU 25, and PHE 29. Harmine, while also demonstrating significant interactions, showed a slightly lower binding affinity of-7.2 kcal/mol and formed interactions with residues such as SER 310, LYS 81, ASP 314, ALA 84, TRP 90, CYS 186, TRP 99, and PHE 92. MD simulations confirmed the stable binding of geissospermine to D1R, as reflected by consistent RMSD values over the simulation period.
Conclusion: Geissospermine’s superior binding affinity and stable interaction with D1R highlight its potential as a therapeutic candidate for neurological disorders, particularly Huntington’s disease, where modulation of dopamine signaling could yield significant clinical benefits. These findings warrant further investigation into the neuroprotective potential of geissospermine in dopaminergic dysfunction.
Keywords: Geissospermine, Harmine, Dopamine D1 Receptors, Molecular dynamic simulation, In silico
© 2025 The Authors. Published by Innovare Academic Sciences Pvt Ltd. This is an open access article under the CC BY license (https://creativecommons.org/licenses/by/4.0/)
DOI: https://dx.doi.org/10.22159/ijap.2025v17i2.51119 Journal homepage: https://innovareacademics.in/journals/index.php/ijap
INTRODUCTION
The Dopamine D1 receptor (D1R) plays a pivotal role in the Central Nervous System (CNS), particularly in modulating neuronal signaling pathways that are critical for various physiological functions, including motor control, cognition, mood regulation, and the reward system [1, 2]. As a G protein-coupled receptor (GPCR), D1R primarily activates the Cyclic Adenosine Monophosphate (cAMP) signaling pathway through the stimulation of adenylyl cyclase, influencing intracellular signaling cascades that modulate neuronal excitability and synaptic plasticity. Given its widespread expression in the striatum, prefrontal cortex, and limbic system, D1R is integral to both normal neurological function and the pathogenesis of several neurodegenerative and psychiatric disorders [3].
Dysregulation of D1R signaling has been implicated in a range of neurological diseases, including Huntington's disease [4], Parkinson's disease [5], schizophrenia [6], and substance abuse disorders [7]. In Huntington's disease, the progressive degeneration of medium spiny neurons in the striatum leads to altered dopaminergic signaling, including impaired D1R function, contributing to motor dysfunction and cognitive decline [4]. Similarly, in Parkinson's disease, the degeneration of dopaminergic neurons in the substantia nigra diminishes dopamine availability, severely disrupting D1R-mediated signaling pathways and resulting in the characteristic motor symptoms of the disease [5]. Beyond neurodegenerative disorders, D1R has been implicated in the etiology of schizophrenia, where an imbalance in dopamine receptor activity, including both D1R and D2R, contributes to cognitive dysfunction and altered reward processing [6]. In the context of substance abuse, D1R is heavily involved in the dopaminergic reward system, influencing addictive behaviours by modulating the brain's response to rewarding stimuli such as drugs of abuse [7]. Given its critical role in these processes, D1R represents a potential therapeutic target for mitigating symptoms or slowing disease progression in a variety of CNS disorders.
The search for effective therapeutic agents that can target D1R while also providing neuroprotective effects is an ongoing challenge in neuropharmacology. While numerous synthetic compounds have been developed to modulate D1R activity, there is a growing interest in exploring naturally derived compounds for their potential neuroprotective properties [8, 9]. Phytocompounds, in particular, have gained attention due to their ability to interact with multiple biological targets, their generally lower toxicity, and their long-standing use in traditional medicine. In this context, the alkaloid compounds geissospermine and harmine have emerged as promising candidates for modulating dopaminergic signaling and offering neuroprotection [10].
Geissospermine, an indole alkaloid derived from plants such as Geissospermumvellosii, has been traditionally used in South American medicine for its analgesic, anti-inflammatory, and antispasmodic properties. Preliminary studies have suggested that geissospermine may possess neuroprotective qualities, potentially through its ability to modulate ion channels and inhibit neuroinflammation—mechanisms that are crucial in preventing or slowing neurodegenerative processes [11, 12]. Harmine, a beta-carboline alkaloid found in Banisteriopsiscaapi and other plants, has been widely studied for its Monoamine Oxidase-A (MAO-A) inhibitory activity, which increases the levels of serotonin, dopamine, and norepinephrine in the brain. Harmine’s ability to enhance neurogenesis, protect neurons from oxidative damage, and inhibit the aggregation of beta-amyloid plaques has led to its consideration as a potential therapeutic agent for neurodegenerative diseases such as Alzheimer’s and Parkinson’s diseases [13, 14].
Given the potential neuroprotective properties of geissospermine and harmine, this study aims to evaluate their interactions with D1R using in silico methods, including molecular docking and dynamic simulations. By assessing their binding affinities and stability in complex with D1R, the study seeks to determine whether these phytocompounds can modulate D1R activity in a manner that may offer therapeutic benefits for neurodegenerative disorders. Additionally, the pharmacokinetic and ADMET (Absorption, Distribution, Metabolism, Excretion, and Toxicity) profiles of these compounds are examined to assess their suitability as drug candidates. Through this investigation, the study aims to contribute to the growing body of research exploring the neuroprotective potential of natural compounds, offering novel insights into the therapeutic applications of geissospermine and harmine in targeting D1R-mediated pathologies.
MATERIALS AND METHODS
Retrieval of ligands
Canonical Simplified Molecular Input Line Entry System (SMILES) representations of this geissospermine and harmine were retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/) for subsequent pharmacological evaluation [15]. Additionally, 2D Structure Data Files (SDF) were acquired for docking studies. The SDF file was imported into PyRx, where it underwent energy minimization using the Universal Force Field (UFF) and conversion to Protein Data Bank (PDB) format, facilitating ligand structure visualization. Ligand preparation for docking was conducted utilizing the PyRx software (https://pyrx.sourceforge.io/) [15].
Retrieval of proteins
The proteins implicated in the pathophysiology of Huntington's disease are identified as potential targets for geissospermine and harminephyto compounds. Consequently, crystal structures of Dopamine D1R (PDB ID: 7JOZ), were retrieved from the RCSB Protein Data Bank (https://www.rcsb.org/). These structures were resolved via X-ray diffraction. Preparation of the protein structures for docking entailed the removal of non-structural elements, including water molecules and chains or heteroatoms, while missing atoms were added. Additionally, polar hydrogen atoms were introduced, and Kollman charges were applied to ensure uniform charge distribution on the protein surface [16].
Structure validation
To verify the structural integrity of the preparations, secondary structural validation was conducted using PDBsum generate (http://www.ebi.ac.uk/thornton-srv/databases/pdbsum/Generate.html). Furthermore, a hydrophilicity plot was generated using DS Biovia Discovery Studio [17] visualizer to assess the hydrophobic or hydrophilic nature of amino acids within the protein.
Molecular docking and visualization
PyRx, a virtual screening software utilized in computer-based drug discovery, enables the screening of compound libraries against potential drug targets. In this study, multiple ligand-protein dockings were performed using AutoDockVina 2.0, integrated into the PyRx software (https://pyrx.sourceforge.io/). The primary objective of molecular docking was to evaluate the inhibitory potential of geissospermine and harminephytocompounds against Dopamine D1R, which are implicated in the pathophysiology of Huntington's disease. The purified proteins served as macromolecules, while the ligands were considered flexible entities for docking. Subsequently, a grid box was generated for all proteins (refer to table 1). Docking results were quantified based on binding affinity, reflecting the energy associated with ligand binding. Throughout the docking process, ligands maintained a total of nine degrees of flexibility, with the binding pose exhibiting zero Root mean Square Deviation (RMSD) deemed most optimal due to maximal binding efficacy. The molecule demonstrating the lowest binding affinity, coupled with favourable ADMET characteristics, was identified as the most promising candidate for anti-pancreatitis therapy. Visualization of docking interactions with the lowest binding energy was facilitated using DS Biovia Discovery Studio [18].
Table 1: Docking grid parameters of the macromolecules
| Macromolecule | PDB ID | Centre dimension | Dimension (Angstrom) | ||||
| X | Y | Z | X | Y | Z | ||
| Dopamine-D1R | 7JOZ | 23.1720 | 1.2113 | 23.6072 | 46.3979 | 36.2038 | 52.6852 |
Pharmacological studies
In assessing the quality of a drug, it is imperative to ensure its efficacy in achieving therapeutic outcomes and its compatibility with appropriate dosage levels, alongside meeting acceptable standards for Absorption, Distribution, Metabolism, and Excretion (ADME) parameters. To address these criteria, the phytocompoundsgeissospermine and harmineunderwent ADME analysis utilizing SwissADME (http://www.swissadme.ch/), while their toxicity endpoints were evaluated using ADMETlab 2.0 (https://admetmesh.scbdd.com/) This aids in elucidating the crucial pharmacological properties and safety profiles of chemical compounds, thereby facilitating informed decision-making throughout the drug development process [19].
Molecular dynamic simulation
Molecular Dynamics Simulation (MDS) conducted using GROMACS software aimed to evaluate the stability of the geissospermine_dopamineprotein complex. Docking analysis revealed a low binding affinity between the phytocompound-geissospermine (PubChem CID: 5281401) and Dopamine D1R. Utilizing the GROMACS package with GROMOS96 54a7 force field, MDS was executed. The geissospermine structure was prepared using the ATB Server. Simulation parameters included a cubic box dimension of X=90, Y=90, and Z=90. Energy minimization and equilibration via the steepest descent method optimized volume. Solvation utilized the SPCE water model, maintaining a 0.15M salt concentration with NA+ and Cl-ions, and the complex's pH was set at 7. Equilibration consisted of two phases: a 10-ns v-rescale algorithm heating period gradually increasing temperature to ~300 K in the NVT ensemble, followed by a 5-ns restrained phase allowing solvent settling. This was succeeded by a 10-ns NPT equilibration phase with gradually removed restraints. Equilibrated systems underwent a 200 ns preparation period for graphing and MMPBSA analysis for the last 50 ns, while maintaining pressure at 1 bar and temperature at 300 K. MDS evaluated variations in RMSD, RMSF, Rg, SASA, and hydrogen bond count of the complex, enabling a comprehensive stability assessment [20].
RESULTS
Retrieval of ligands
The canonical SMILES and the 2D SDF structures of the geissospermine and harmine identified from IMPPAT and were obtained from PubChem. Subsequently, both the phytocompounds underwent pharmacological profiling and docking analysis. The structure of the top phytocompound geissospermine (PubChem CID: 5281401) is illustrated in fig. 1.
Structure validation
The structures of Dopamine D1R(PDB ID: 7JOZ), were subjected to secondary structure validation (fig. 2). The purified Dopamine D1R structure (fig. 2a) comprises 454 amino acids, including 1 sheet, 1 beta-hairpin, 1 betabulge, 2 strands, 12 helices, 22 helix-helix interactions, and 14 beta turns as secondary structural elements (fig. 2b). Notably, 93.4% (171 amino acids) of the residues were located in the most favored region of the Ramachandran Plot (fig. 2d). Dopamine D1R, like other nuclear receptors, exhibits a hydrophobic nature (fig. 2c).
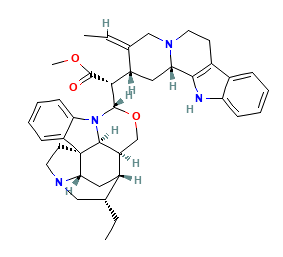
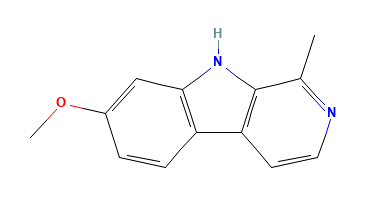
Fig. 1: 2-dimensional structures of geissospermine and harmine
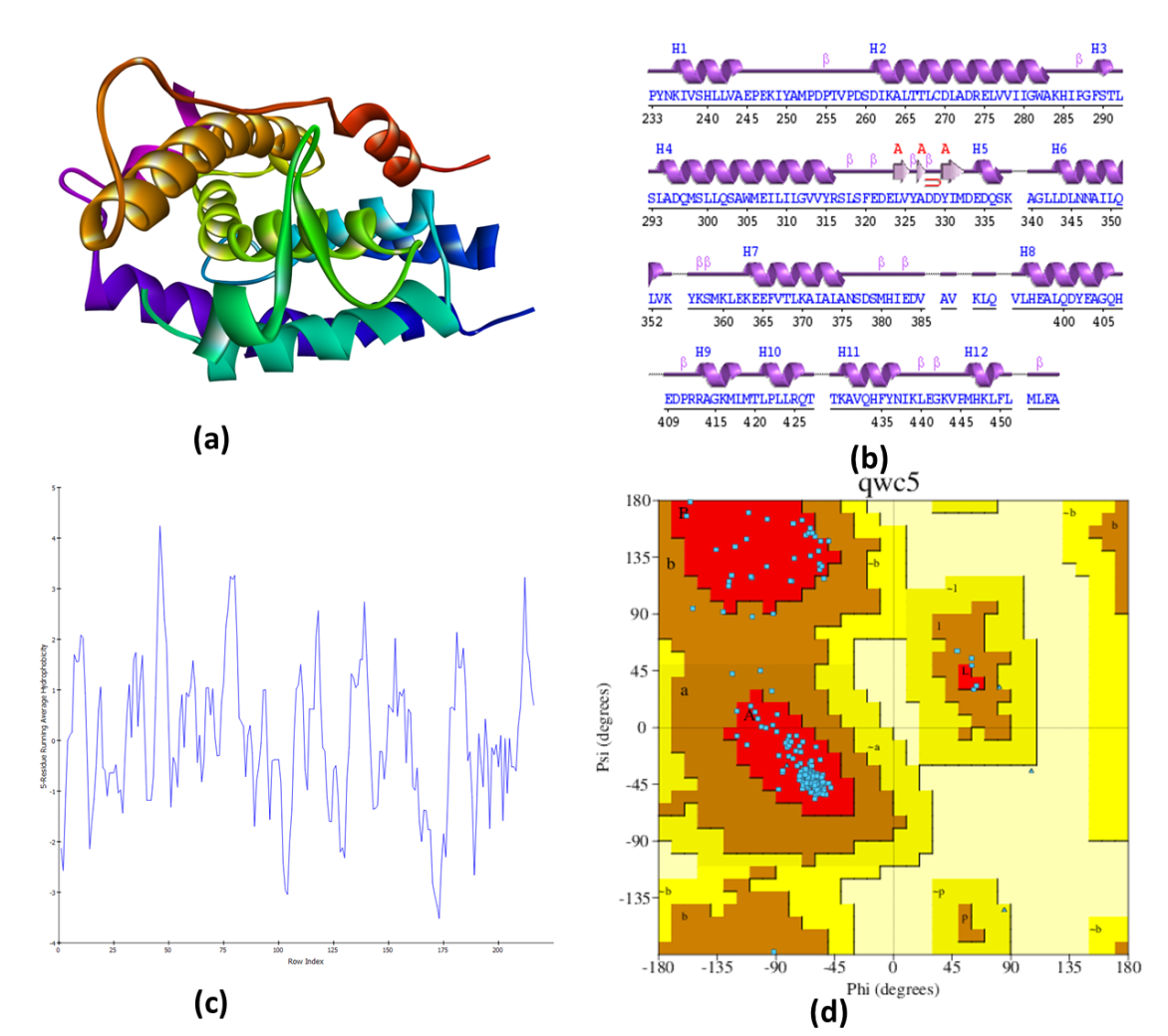
Fig. 2: Structure validation of dopamine D1R (PDB ID: 7JOZ), 3D structure (b) Secondary structure wiring diagram (c)Hydropathy plots (d) Ramachandran plot
Molecular docking and visualization
The investigation into the molecular docking results of geissospermine and harmine (fig. 4) phytocompounds, as outlined in table 2, revealed varying degrees of binding affinity with selected target receptors. Among the three receptors analyzed, the phytocompounds demonstrated the least binding affinity towards the Dopamine D1R.
Of particular interest in this study is the phytocompound geissospermine (PubChem CID: 5281401), which exhibited notably low binding affinity towards the Dopamine D1R protein, with a recorded value of-8.6 kcal/mol. This value represents the most favourable binding affinity among all tested phytocompounds against the Dopamine D1R protein. Additionally, geissospermine (PubChem CID: 5281401) consistently displayed low binding affinities across all three receptors, thus positioning it as a promising candidate.
Table 2: Molecular docking of geissospermine with target receptors
| Ligand | Dopamine D1R (Kcal/mol) |
| Geissospermine | -8.6 |
| Harmine | -7.2 |
In the molecular docking of phytocompound geissospermine (PubChem CID: 5281401) with Dopamine D1R (PDB ID: 7JOZ) the amino acids including ILE 308, LYS 370, VAL 278, ILE 249, LYS 248, TYR 315 and ARG 316 were predominantly interacting with geissospermine (fig. 3).
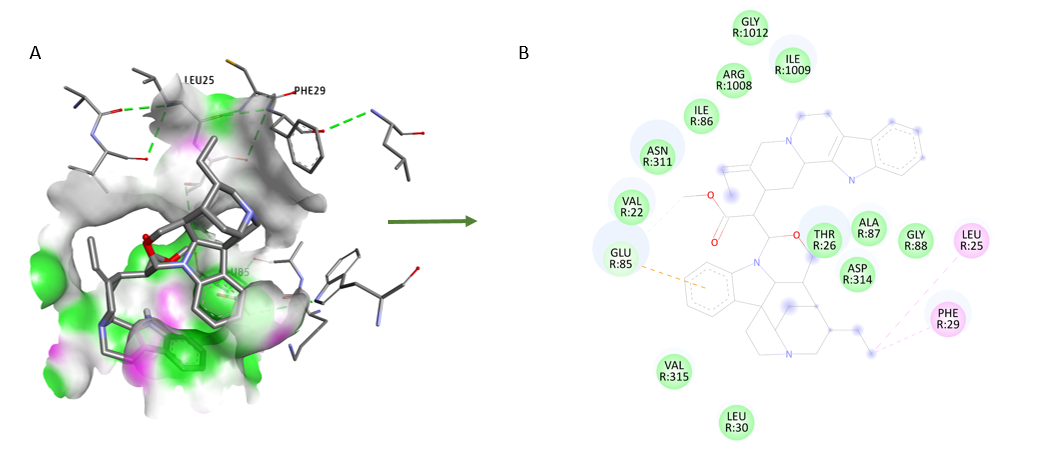
Fig. 3: Molecular docking visualization of phytocompound geissospermine (PubChem CID: 5281401) with Dopamine D1R (PDB ID: 7JOZ)
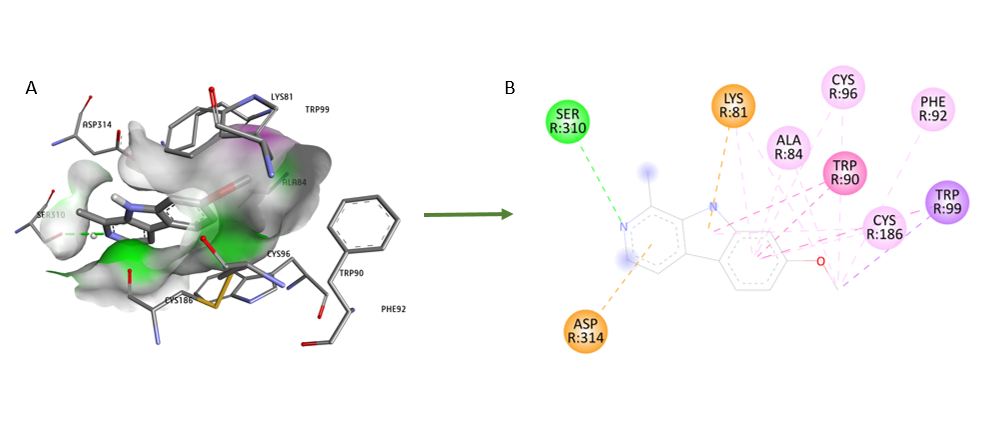
Fig. 4: Molecular docking visualization of phytocompound harmine (PubChem CID: 5281401) with dopamine D1R (PDB ID: 7JOZ)
Pharmacological studies
The compound geissospermine (PubChem CID: 5281401) underwent an ADMET evaluation following its selection as the most promising candidate from molecular docking studies. Through in silico pharmacological profiling, it was observed that geissospermine possesses favorable physicochemical properties. Notably, the compound demonstrates solubility, supported by an optimal number of hydrogen bond donors and acceptors, which are crucial determinants of drug-target interactions, contributing to binding affinity and selectivity. Additionally, these hydrogen bonds play a role in enhancing the compound's solubility in aqueous and polar solvents (table 3).
Geissospermine meets the criteria outlined by Lipinski, Pfizer Rule, and the Golden Triangle screening, indicating its suitability for drug-like properties. Pharmacokinetic analysis indicates that geissospermine does not serve as a substrate for P-glycoproteins (P-gp's), suggesting its potential to inhibit P-gp activity and thereby increase drug bioavailability and tissue distribution, consequently enhancing its therapeutic efficacy. Similarly, cytochromes, a family of enzymes crucial for drug metabolism, were found to metabolize the ligand, facilitating its elimination from the body. Assessments, including Brenk, PAINS, BMS Rule, and Chelator Rule, did not reveal any chemical substructures or functional groups in geissospermine associated with nonspecific or promiscuous interactions with biological targets. Moreover, the ligand exhibited favourable toxicity parameters, with a synthetic accessibility score of 4.88, indicating its ease of synthesis (table 3).
However, it is noteworthy that despite the favourable pharmacological properties outlined above, geissospermine (PubChem CID: 5281401) demonstrated lower gastrointestinal (GI) absorption in preliminary assessments. While optimal GI absorption is typically preferred for oral drug candidates, it is essential to recognize that the compound's exceptional physicochemical properties, drug-like characteristics, and absence of undesirable chemical substructures contribute significantly to its overall potential as a therapeutic agent (table 3).
Molecular dynamic simulation
The primary protein-ligand complex under study, geissospermine complex, was subjected to molecular dynamics simulations using gromacs-2019.4 to explore its internal stability. The ligand topology was obtained from the ATB server for force field coordinates. Each equilibrated structure within the NPT ensemble underwent a 100 ns production run. Trajectory analysis was performed using GROMACS to evaluate various parameters, including Protein RMSD (Root mean Square Deviation), RMSF (Root mean Square Fluctuation), RG (Radius of Gyration), SASA (Solvent Accessible Surface Area), and H-Bond parameters. The "Average RMSD" and "Average RMSF" metrics are utilized as indicators of the protein's overall stability and flexibility, respectively (table 4). Lower RMSD values suggest greater structural stability, while higher RMSF values indicate increased flexibility within the protein structure. The "Radius of Gyration" quantifies the protein's size, with larger values indicating a larger protein structure. The "Average SASA" measures the extent of the protein surface exposed to the solvent, where higher values suggest more extensive solvent exposure. Energy values obtained from the simulations are documented in table 4, fig. 5-7.
Table 3: Pharmacological profiling of geissospermine (PubChem CID: 5281401)
| Physicochemical properties | |||||||||
| Formula | MW | Heavy atoms | Fraction Csp3 | Rot bonds | HA | HD | MR | TPSA | nRing |
| C25H22O10 | 482.44g/mol | 25 | 0.24 | 4 | 10 | 6 | 121.05 | 166.14Ų | 5 |
| Pharmacokinetics | |||||||||
| Lipophilicity | Water solubility | GIA | BBB | P-gp substrate | CYP1A2 inhibitor | CYP2C19 inhibitor | CYP2C9 inhibitor | CYP2D6 inhibitor | CYP3A4 inhibitor |
| 1.56 | Soluble | Low | No | No | No | No | No | No | Yes |
| Drug-likeness | |||||||||
| Lipinski | Pfizer rule | Golden triangle |
GSK rule | BA | Brenk | PAINS | BMS rule | Chelator rule | SA |
| Yes | Accepted | Accepted | Rejected | 0.55 | No | No | No | No | 4.88 |
| Toxicity | |||||||||
| hERG blockers | H-HT | DILI | AMES | ROAT | Carcino genecity | Eye corrosion | RT | ATC | GCR |
| Inactive | Inactive | Active | Inactive | Moderate | Inactive | Inactive | Inactive | No Alerts | |
MW: Molecular weight (<500 g/MOL); Csp3:>0.25 and<1; Rot bonds: Rotatable bonds(0-1); HA: Hydrogen bond acceptors (less than 10); HD: Hydrogen bond donors(less than 5): MR: Molar refractivity (40-130); TPSA: Topological surface area (20-130 Ų); nRing: Number of Rings (0-6); Lipophilicity:>-0.7 and<5.0: GIA: Gastrointestinal absorption; BBB: Blood-brain barrier permeation; Pgp: Permeability glycoproteins: BA: Bioavailability: PAINS: Pan Assay Interference Compounds; SA: Synthetic Accessibility (1-10); H-HT: Human Hepatotoxicity; DILI: Drug-Induced Liver Injury; ROAT: Rat Oral Acute Toxicity; RT: Respiratory Toxicity; ATC: Acute Toxicity Rule; GCR: Genotoxic Carcinogenicity Rule
Table 4: Molecular dynamics of geissospermine-dopamine D1R complex
| Protein | Average RMSD (nm) | RMSF (nm) | Radius of gyration (nm) | Average SASA (nm2) |
| Geissospermine-dopamine D1R complex | 0.21+/-0.01 | 0.14+/-0.05 | 1.47+/-0.02 | 82.46+/-2.17 |
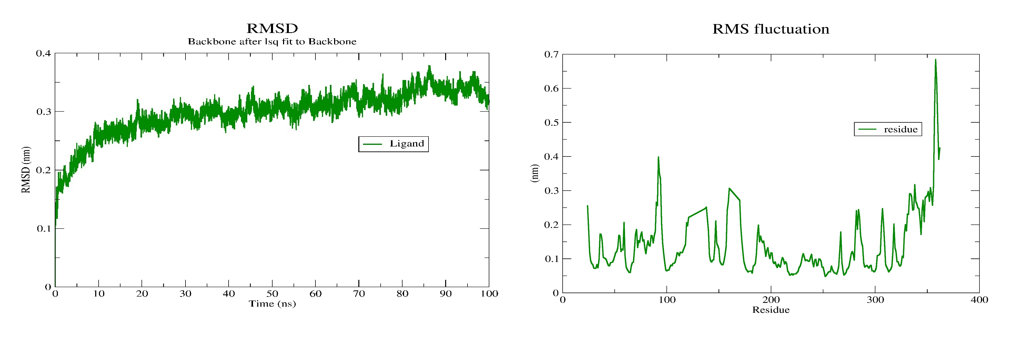
Fig. 5: RMSD plot and RMSF plot
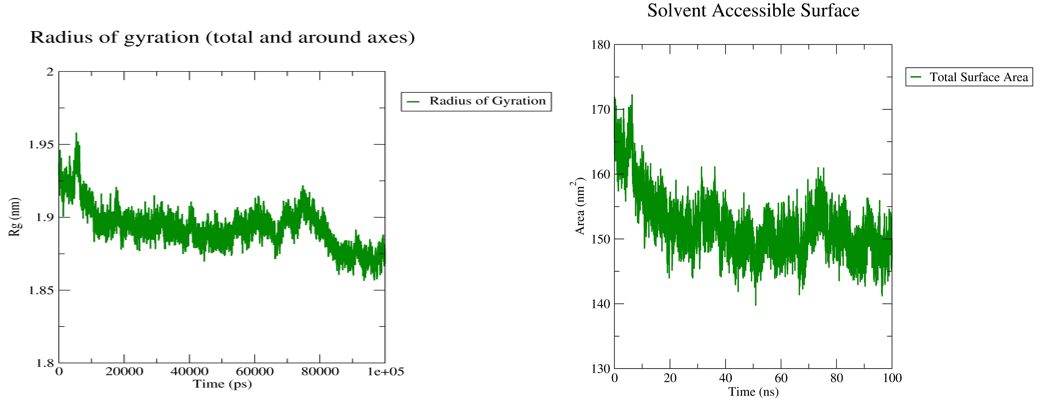
Fig. 6: Radius of gyration plot and SASA plot
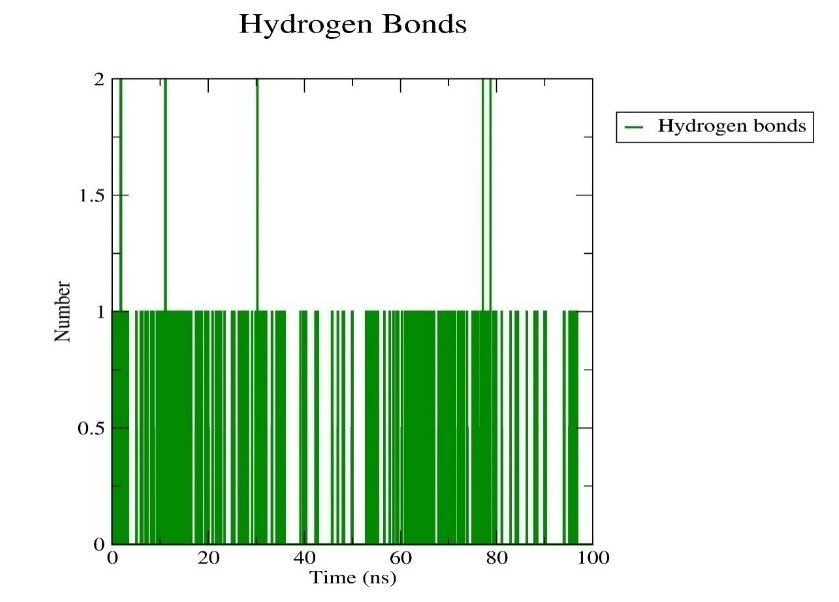
Fig. 7: H-bond plot
DISCUSSION
The Dopamine D1 receptor (D1R) is a significant member of the dopamine receptor family, which plays a crucial role in the central nervous system's functioning. D1R is a G protein-coupled receptor (GPCR) that is primarily involved in regulating dopaminergic signaling pathways. Dopaminergic transmission is essential for controlling various brain functions, including motor control, cognition, emotion, and reward pathways [21]. The D1R protein is primarily located in the striatum, frontal cortex, and limbic areas of the brain, where it mediates the activation of adenylyl cyclase, leading to the production of Cyclic Adenosine Monophosphate (cAMP). This signaling cascade is essential for modulating neuronal activity and synaptic plasticity [1]. The dysregulation of D1R is implicated in several neurodegenerative and psychiatric disorders, including Parkinson's disease, Huntington's disease, schizophrenia, and substance abuse disorders [4-7].
D1R is considered a major biomarker in neurodegenerative diseases due to its significant involvement in dopaminergic pathways that are central to these diseases' pathogenesis. In Huntington’s disease, for instance, the progressive loss of medium spiny neurons in the striatum results in impaired dopamine signaling through D1R, which exacerbates motor dysfunction and cognitive decline [2]. In Parkinson’s disease, the death of dopaminergic neurons in the substantia nigra leads to a loss of dopamine transmission to D1R-expressing neurons, resulting in motor impairments such as bradykinesia, rigidity, and tremor [5]. As such, D1R not only plays a crucial role in the pathogenesis of these diseases but also serves as a potential target for therapeutic interventions aimed at modulating dopamine signaling to mitigate symptoms or slow disease progression. Given its central role in dopamine-related neurodegeneration, D1R is a valuable target for neuroprotective agents that aim to restore or modulate dopamine signaling in affected neurons [22].
In this study, the phytocompounds geissospermine and harmine were selected for their potential neuroprotective effects on the D1R protein. Geissospermine, an alkaloid derived from plants such as Geissospermumvellosii, and harmine, a beta-carboline alkaloid found in Banisteriopsiscaapi, have both been traditionally used in ethnomedicine for their neurological benefits. Geissospermine has been shown in various studies to possess antispasmodic, analgesic, and sedative properties [11]. Its neuroprotective potential has been suggested in studies exploring its ability to modulate ion channels and inhibit neuroinflammation, which are key contributors to neurodegenerative diseases [12]. Harmine, on the other hand, is well-known for its Monoamine Oxidase A (MAO-A) inhibitory properties, which enhance the levels of serotonin, dopamine, and norepinephrine in the brain [13]. This has led to harmine being explored for its antidepressant effects, as well as its neuroprotective potential in Parkinson's and Alzheimer's disease models. Studies have shown that harmine can promote neurogenesis, protect neurons from oxidative stress, and inhibit the formation of beta-amyloid plaques, which are hallmarks of Alzheimer's disease [14]. The neuroprotective properties of both geissospermine and harmine make them promising candidates for modulating dopaminergic signaling through D1R, especially in neurodegenerative conditions such as Huntington’s and Parkinson’s diseases. Natural compounds like geissospermine and harmine offer several advantages over synthetic drugs due to their ability to interact with multiple biological targets, lower toxicity, and better biocompatibility. These characteristics make phytocompounds appealing candidates for drug development, especially in the context of chronic neurodegenerative diseases, where long-term safety is a primary concern.
The aim of this study was to evaluate the neuroprotective potential of geissospermine and harmine on the D1R protein using in silico techniques, including molecular docking, pharmacological profiling, and molecular dynamic simulations. The study sought to determine the binding affinity of these compounds to D1R and explore their pharmacokinetic properties, stability, and interactions with the receptor. The molecular docking results revealed that geissospermine exhibited a lower binding affinity (-8.6 kcal/mol) compared to harmine (-7.2 kcal/mol), suggesting that geissospermine may form more stable and favorable interactions with D1R. The docking analysis also highlighted the involvement of key amino acid residues, including ILE 308, LYS 370, VAL 278, ILE 249, and TYR 315, which interact with geissospermine within the D1R binding pocket. These interactions, particularly the hydrogen bonds formed with TYR 315 and ARG 316, may contribute to the higher binding affinity observed for geissospermine, indicating its potential as a more effective neuroprotective agent for targeting D1R.
Molecular docking revealed that geissospermine interacts more favorably with key residues in the D1R binding pocket, which could indicate stronger inhibitory potential or better modulation of receptor activity. This result is significant because it suggests that geissospermine can effectively target critical residues involved in dopamine signaling, which is essential in the context of neurodegenerative diseases. In contrast, while harmine also demonstrated significant binding, its lower affinity suggests it might not be as robust as geissospermine in modulating D1R activity. Nonetheless, both compounds exhibited interactions with important residues, which supports their role as neuroprotective agents.
The pharmacological profiling of geissospermine revealed favorable physicochemical properties, including solubility, compliance with Lipinski’s rule, and no significant interactions with P-glycoproteins, suggesting its suitability as a drug candidate. The ADMET analysis further showed that geissospermine does not inhibit major cytochrome enzymes, reducing the likelihood of adverse drug-drug interactions. However, the compound demonstrated low gastrointestinal absorption, which may limit its effectiveness as an oral drug. Despite this, the synthetic accessibility and lack of toxic chemical substructures enhance its potential for further drug development. Harmine, while demonstrating lower binding affinity, also exhibited favorable ADMET properties, including good solubility and low toxicity, supporting its continued exploration as a neuroprotective agent.
The molecular dynamic simulations provided valuable insights into the stability of the geissospermine-D1R complex. The low RMSD values (0.21±0.01 nm) indicated that the complex remains structurally stable throughout the simulation, with minimal deviations from the initial docked position. This stability is crucial for ensuring that the compound maintains its binding efficacy over time. The RMSD graph demonstrated that after an initial period of fluctuations, the complex stabilized, maintaining consistent structural integrity throughout the 100 ns simulation. This indicates that geissospermine forms a strong and stable interaction with D1R, which is promising for its long-term efficacy as a neuroprotective agent. The RMSF analysis showed that the flexibility of individual residues within the protein remained relatively low, with some fluctuations observed in specific regions, suggesting that geissospermine does not induce significant conformational changes in the receptor. This is beneficial for drug-target interaction stability, as lower flexibility often correlates with more stable binding. Furthermore, the consistent formation of hydrogen bonds further supports the stability of the ligand-receptor complex, which is essential for long-term neuroprotection. The hydrogen bond analysis revealed that geissospermine formed stable bonds with critical residues throughout the simulation, further strengthening the case for its neuroprotective potential.
The Radius of gyration (Rg) and Solvent-Accessible Surface Area (SASA) analyses indicated that the protein maintained a compact structure with limited solvent exposure, reinforcing the notion that geissospermine is well-embedded within the D1R binding pocket. The SASA values showed a decrease over time, indicating that the ligand became more deeply embedded in the receptor, thereby shielding critical interactions from the solvent environment. The decrease in solvent exposure suggests that geissospermine is likely to remain protected within the binding pocket, reducing the chances of disruption by external factors. The stability of this interaction, as evidenced by both Rg and SASA results, further supports the conclusion that geissospermine is a potent neuroprotective agent capable of maintaining strong and stable interactions with D1R.
The results of this study are significant as they provide novel insights into the potential of geissospermine as a neuroprotective agent targeting D1R. The combination of favorable binding affinity, pharmacokinetic properties, and molecular stability suggests that geissospermine could be developed as a therapeutic agent for neurodegenerative diseases characterized by D1R dysregulation, such as Huntington’s and Parkinson’s diseases. Harmine, while less potent in binding to D1R, still offers valuable neuroprotective properties and may be considered in combination therapies to enhance its efficacy.
The primary limitation of this study lies in its in-silico nature. While molecular docking and dynamic simulations provide valuable insights into the potential interactions between ligands and proteins, experimental validation is necessary to confirm these findings. Further in vitro and in vivo studies are required to evaluate the neuroprotective effects of geissospermine and harmine in animal models of neurodegenerative diseases. Additionally, the low gastrointestinal absorption of geissospermine presents a challenge for oral administration, and future studies should explore alternative delivery methods, such as intranasal or transdermal routes, to improve its bioavailability.
This study identifies geissospermine as a promising neuroprotective agent for targeting the Dopamine D1 receptor in neurodegenerative diseases. The favorable binding affinity, structural stability, and pharmacological properties of geissospermine highlight its potential for further drug development. Harmine also demonstrates neuroprotective properties, albeit with lower efficacy in targeting D1R. Future research should focus on optimizing the pharmacokinetic properties of geissospermine, exploring its effects in animal models, and investigating its potential in combination therapies. The results of this study lay the groundwork for developing geissospermine as a novel therapeutic agent for neurodegenerative diseases, with the potential to modulate dopaminergic signaling and protect neurons from degeneration.
CONCLUSION
This study provides valuable insights into the neuroprotective potential of geissospermine and harmine through their interactions with the Dopamine D1 receptor (D1R) as assessed via molecular docking and molecular dynamics simulations. Geissospermine demonstrated a notably higher binding affinity towards D1R compared to harmine, likely due to stronger and more stable interactions within the receptor's binding pocket. The molecular dynamics simulations confirmed the stability of this interaction over time, reinforcing geissospermine’s promise as a potential therapeutic candidate for treating neurological disorders involving dopaminergic dysfunction. These findings not only enhance our understanding of natural compounds like geissospermine as modulators of D1R activity but also lay the groundwork for future experimental studies and drug development efforts aimed at harnessing the neuroprotective properties of these phytocompounds.
FUNDING
Nil
AUTHORS CONTRIBUTIONS
Sriranjini A. S: Conceptualization, Methodology, Data Curation, Formal Analysis, Critical Evaluation, Visualization, Writing – Original Draft, Dr. Ashish Thapliyal: Supervision, Validation, Critical Evaluation, Writing – Review and Editing, Project Administration, Dr. Kumud Pant: Literature Review, Resources, Critical Evaluation, Writing – Review and Editing.
CONFLICT OF INTERESTS
Declared none
REFERENCES
Possemato E, LA Barbera L, Nobili A, Krashia P, D Amelio M. The role of dopamine in NLRP3 inflammasome inhibition: implications for neurodegenerative diseases. Ageing Res Rev. 2023;87:101907. doi: 10.1016/j.arr.2023.101907, PMID 36893920.
HE J, Zhu G, Wang G, Zhang F. Oxidative stress and neuroinflammation potentiate each other to promote progression of dopamine neurodegeneration. Oxid Med Cell Longev. 2020 Jul 3;2020:6137521. doi: 10.1155/2020/6137521, PMID 32714488, PMCID PMC7354668.
LI H, Yang P, Knight W, Guo Y, Perlmutter JS, Benzinger TL. The interactions of dopamine and oxidative damage in the striatum of patients with neurodegenerative diseases. J Neurochem. 2020;152(2):235-51. doi: 10.1111/jnc.14898, PMID 31613384, PMCID PMC6981021.
Moreno Delgado D, Puigdellivol M, Moreno E, Rodriguez Ruiz M, Botta J, Gasperini P. Modulation of dopamine D1 receptors via histamine H3 receptors is a novel therapeutic target for huntingtons disease. Elife. 2020;9:e51093. doi: 10.7554/eLife.51093, PMID 32513388, PMCID PMC7282811.
Kaasinen V, Vahlberg T, Stoessl AJ, Strafella AP, Antonini A. Dopamine receptors in parkinsons disease: a meta-analysis of imaging studies. Mov Disord. 2021;36(8):1781-91. doi: 10.1002/mds.28632, PMID 33955044.
Martel JC, Gatti MC Arthur S. Dopamine receptor subtypes physiology and pharmacology: new ligands and concepts in schizophrenia. Front Pharmacol. 2020;11:1003. doi: 10.3389/fphar.2020.01003, PMID 32765257, PMCID PMC7379027.
Botticelli L, Micioni DI, Bonaventura E, Del Bello F, Giorgioni G, Piergentili A, Romano A. Underlying susceptibility to eating disorders and drug abuse: genetic and pharmacological aspects of dopamine D4 receptors. Nutrients. 2020;12(8):2288. doi: 10.3390/nu12082288, PMID 32751662, PMCID PMC7468707.
DE Bartolomeis A, Barone A, Begni V, Riva MA. Present and future antipsychotic drugs: a systematic review of the putative mechanisms of action for efficacy and a critical appraisal under a translational perspective. Pharmacol Res. 2022;176:106078. doi: 10.1016/j.phrs.2022.106078, PMID 35026403.
Newman AH, KU T, Jordan CJ, Bonifazi A, XI ZX. New drugs old targets: tweaking the dopamine system to treat psychostimulant use disorders. Annu Rev Pharmacol Toxicol. 2021;61(1):609-28. doi: 10.1146/annurev-pharmtox-030220-124205, PMID 33411583, PMCID PMC9341034.
Ang JJ, Low BS, Wong PF. Dopamine and central dopaminergic circuitry in neurodegenerative diseases: roles and mechanisms of action of natural phytochemicals. Pharmacological Research Natural Products. 2024 Jun;3:100050. doi: 10.1016/j.prenap.2024.100050.
Lima JA, Costa R TW, DA Fonseca ACC, DO Amaral RF, Nascimento M DO Santos, DSB Filho OA DE Miranda ALP, Ferreira Neto DC, Lima FRS, Hamerski L, Tinoco LW. Geissoschizoline a promising alkaloid for alzheimers disease: inhibition of human cholinesterases anti-inflammatory effects and molecular docking. Bioorg Chem. 2020 Nov;104:104215. doi: 10.1016/j.bioorg.2020.104215, PMID 32920358.
Lima JA, Costa TW, Silva LL, Miranda AL, Pinto AC. Antinociceptive and anti-inflammatory effects of a geissospermum vellosii stem bark fraction. An Acad Bras Cienc. 2016;88(1):237-48. doi: 10.1590/0001-3765201520140374, PMID 26840005.
Xie Z, Liu W, Dang R, HU X, Cai F, Xiang Z. Effects and mechanisms of harmine on ameliorating ethanol-induced memory impairment. J Ethnopharmacol. 2025;337(1):118789. doi: 10.1016/j.jep.2024.118789, PMID 39241971.
Kadyan P, Singh L. Unraveling the mechanistic interplay of mediators orchestrating the neuroprotective potential of harmine. Pharmacol Rep. 2024;76(4):665-78. doi: 10.1007/s43440-024-00602-8, PMID 38758470.
Jayasurya, Swathy, Susha, Sharma S. Molecular docking and investigation of boswellia serrata phytocompounds as cancer therapeutics to target growth factor receptors: an in silico approach. Int J App Pharm. 2023;15(4):173-83. doi: 10.22159/ijap.2023v15i4.47833.
Vora D, Kapadia H, Dinesh S, Sharma S, Manjegowda DS. Gymnema sylvestre as a potential therapeutic agent for PCOS: insights from mRNA differential gene expression and molecular docking analysis. Futur J Pharm Sci. 2023 Aug;9(1). doi: 10.1186/s43094-023-00529-6.
Bhor S, Tonny SH, Dinesh S, Sharma S. Computational screening of damaging nsSNPs in human SOD1 genes associated with amyotrophic lateral sclerosis identifies destabilising effects of G38R and G42D mutations through in silico evaluation. In Silico Pharmacol. 2024;12(1):20. doi: 10.1007/s40203-024-00191-7, PMID 38559706, PMCID PMC10973320.
Sunkadakatte Venugopal B, Dinesh S, Sharma S, Govinda Srinivasan S, Sosalagere Manjegowda D. Exploring the therapeutic potential of green tea phytocompounds for pancreatic cancer: an mRNA differential gene expression and pathway analysis study. Biomedicine. 2023;43(4):1231-8. doi: 10.51248/.v43i4.2869.
Galgale S, Zainab R, Kumar A. Molecular docking and dynamic simulation-based screening identifies inhibitors of targeted SARS-CoV-2 3clpro and human ace2. Int J App Pharm. 2023;15(6):297-308. doi: 10.22159/ijap.2023v15i6.48782.
Bhowmik R, Nath R, Sharma S, Roy R, Biswas R. High throughput screening and dynamic studies of selected compounds against SARS-CoV-2. Int J App Pharm. 2022;14(1):251-60. doi: 10.22159/ijap.2022v14i1.43105.
Jones Tabah J, Mohammad H, Paulus EG, Clarke PB, Hebert TE. The signaling and pharmacology of the dopamine D1 receptor. Front Cell Neurosci. 2021;15:806618. doi: 10.3389/fncel.2021.806618, PMID 35110997, PMCID PMC8801442.
Yang P, Knight WC, LI H, Guo Y, Perlmutter JS, Benzinger TL. Dopamine D1 + D3 receptor density may correlate with parkinson disease clinical features. Ann Clin Transl Neurol. 2021;8(1):224-37. doi: 10.1002/acn3.51274, PMID 33348472.